































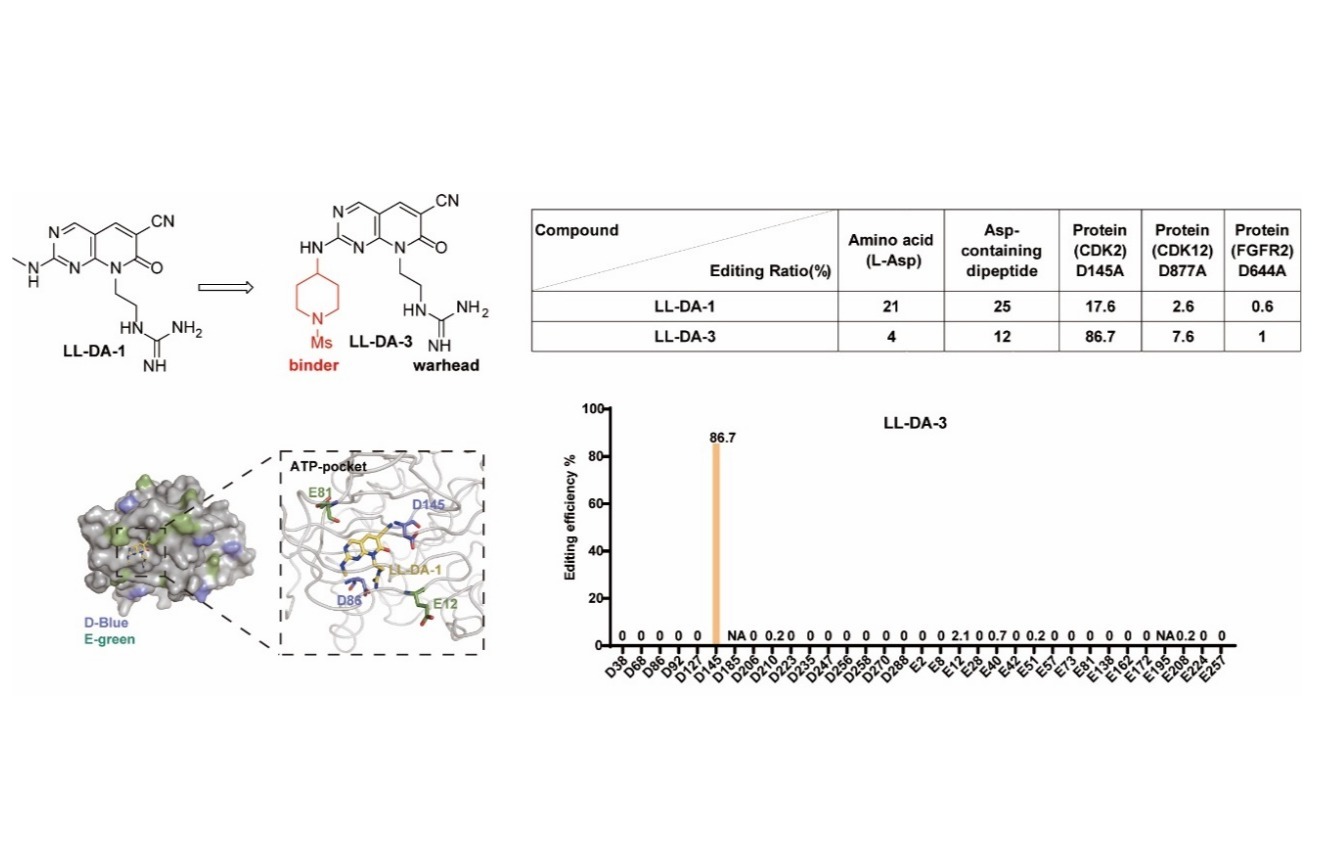
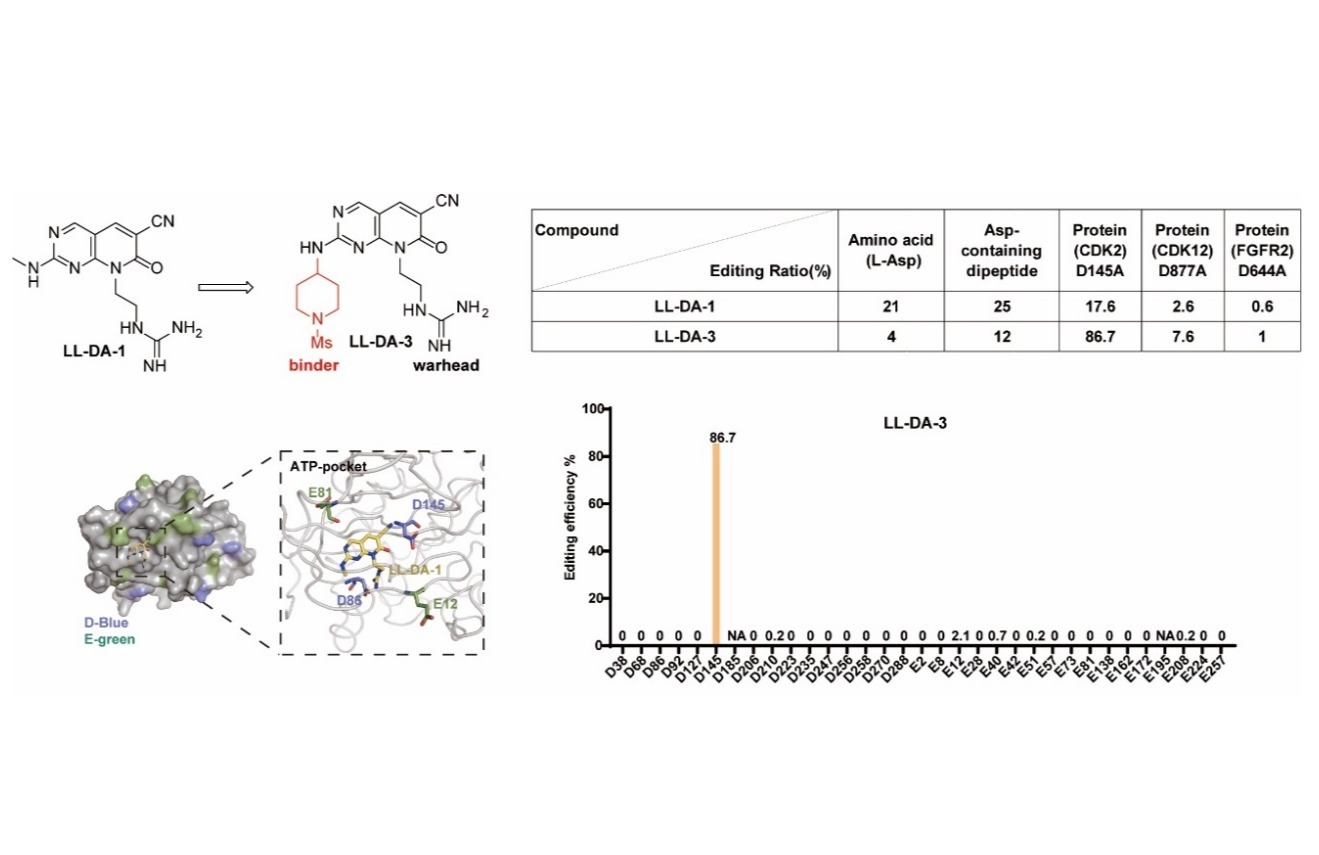

































































































































































































IMMUNITY | 上海药物所揭示K63-polyUb介导的MDA5-MAVS抗病毒信号通路组装程序与激活机制
MDA5是细胞内的异体RNA监测蛋白,属于RIG-I样受体家族(RLRs)的重要成员。MDA5主要参与调控冠状病毒、小核糖核酸病毒、黄病毒和丁型肝炎病毒等RNA病毒引起的免疫反应,是天然免疫的一道重要屏障。MDA5在自身免疫疾病中可识别内源性alu RNA和未经ADAR编辑过的,以及线粒体释放出来的双链RNA。RLRs家族共有RIG-I、MDA5及LGP2三个成员,其中RIG-I和MDA5的N端均拥有串联CARDs结构域。CARDs属于死亡结构域(Death Domain)超家族,可通过CARD-CARD同型相互作用招募MAVS,最终促进I型干扰素(IFN)通路的激活。
泛素化是真核生物中一种重要的翻译后修饰。泛素化可由E1、E2和E3酶及DUBs可逆调节。泛素可以被同型泛素分子修饰并组装成多聚泛素链,包括M1、K11、K27、K48及K63等不同连接的多聚泛素链。其中,K63连接的多聚泛素链(K63-polyUb)在调节RLRs信号通路中起着关键作用[1]。前期研究发现,短链K63-polyUb可以通过共价锚定和非共价锚定两种方式有效地促使RIG-ICARDs的寡聚[2,3]。形成的异源四聚体复合物(K63-polyUb-RIG-ICARDs)可激活MAVSCARD寡聚,形成MAVS纤维的核心[2,3]。然而,K63-polyUb是如何调控MDA5 CARDs组装以及招募、激活MAVS CARD的分子机制,仍是待解决的科学问题。尽管K63-polyUb是MDA5介导的MAVS激活所需要的[4],但目前尚未发现被K63-polyUb共价修饰的MDA5 CARDs。这暗示着非锚定的K63-polyUb在MDA5-MAVS信号传递中的重要作用。
2021年10月12日,中国科学院上海药物研究所郑杰团队在Immunity杂志上以Research Article形式在线发表了题为“Ordered assembly of the cytosolic RNA-sensing MDA5-MAVS signaling complex via binding to unanchored K63-linked poly-ubiquitin chains”的研究成果,揭示了长链,非锚定K63-polyUb促进MDA5-MAVS组装程序与信号传递的分子机制。

首先,研究人员建立了K63-,K48-连接泛素链的生化合成平台,并制备了不同长度的K63-polyUbn(2≤n≤14)(图1)。通过氢氘交换质谱技术(Hydrogen/Deuterium Exchange Mass Spectrometry,HDX-MS),研究人员发现MDA5CARDs和RIG-ICARDs的氢氘交换保护程度依赖于不同长度的K63-polyUbn(MDA5: n≥8; RIG-I: n≥3)而不依赖于K48-polyUbn(n≥10);并且保护强度随着K63-polyUb的长度增加而特异性加强。
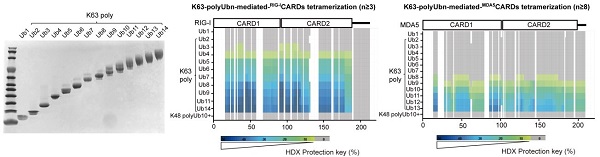
图1:HDX-MS分析K63-polyUb(2≤n≤14)对RLR CARDs寡聚的影响
为了研究K63-polyUbn介导的MDA5CARDs寡聚体的组装机制,研究人员利用冷冻电镜首次解析得到了分辨率为3.3埃 的MDA5CARDs与K63-polyUb13复合体的结构。这也是MDA5CARDs第一个近原子分辨率的冷冻电镜结构。八个Ub分子包围着一个左手螺旋的、环状的CARDs四聚体,其中包括五个Ub-CARD独特的互作界面和七个CARD1-2分子内和分子间独特的互作界面(Ia:Ib,IIa:IIb,和IIIa:IIIb)(图2)。那么MDA5CARDs-K63-polyUbn异源四聚体又是如何招募其下游信号蛋白MAVS?研究人员进一步通过Cryo-EM解析得到了分辨率为3.2埃的由长链K63-polyUb11拴系的“自下而上”的左手螺旋MDA5CARDs-MAVSCARD复合体,MAVSCARD四聚体通过MDA5CARD2-MAVSCARD同型互作的方式堆叠在由八个Ub分子拴绳的MDA5CARDs四聚体顶部。MDA5CARD2与MAVSCARD表位包括三个独特的分子间同型界面(Ia:Ib,IIa:IIb,和IIIa:IIIb)。互作界面氨基酸位点的突变会导致细胞内IFN信号通路激活的缺陷。
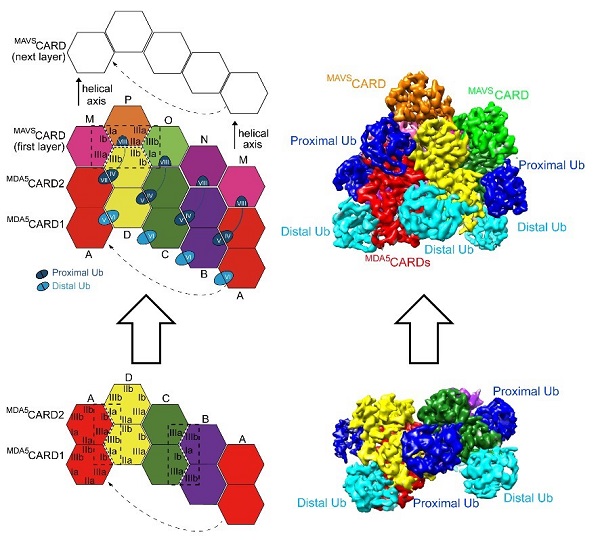
图2:单颗粒冷冻电镜显示K63-polyUbn (n≥8)介导的MDA5-MAVS CARD-CARD的组装机制
同时,研究人员通过生物大分子氢氘交换质谱技术,首次证明了人类MDA5全长蛋白的CARDs在初始状态下处于张开的构象并可与长链K63-polyUb10结合。然而在早期研究中,氢氘交换质谱已经证明了RIG-ICARDs在初始状态下呈闭合的构象[5,6]。这也直接证明了RIG-I和MDA5的CARDs在溶液状态下构象上的巨大差异。其次,研究人员进一步发现K63-polyUb10拴系的MDA5CARDs复合物在溶液中的稳定性受MDA5的RNA依赖的ATP酶活性别构调节:受RNA刺激的MDA5可以结合并水解ATP,其结合产生的能量可以远程地稳定K63-polyUb10-MDA5CARDs的组装以持续传递下游激活信号;相反,未受RNA刺激的MDA5虽然也可以与长链K63-polyUb10结合,但ATP可以使其解聚并防止MDA5不必要的激活(图3)。这些证据表明,在含有高浓度ATP的线粒体膜附近,MDA5对RNA的有效识别有助于远程调控K63-polyUb10-MDA5CARDs复合体的稳定性,并帮助激活MAVS。
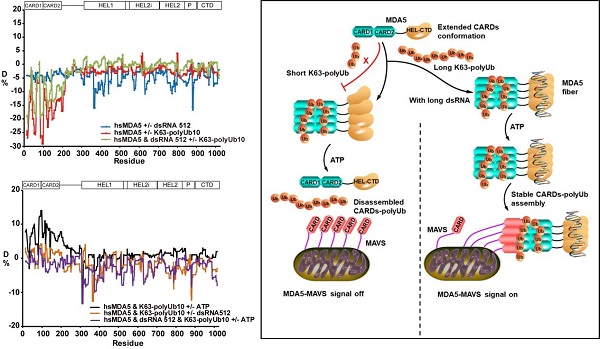
图3:HDX-MS分析全长MDA5在其识别配体或底物作用下(dsRNA/ATP/K63-polyUb)的动态的构象变化与信号传导机制
综上所述,该研究通过生物大分子氢氘交换质谱和冷冻电镜技术发现长链,非锚定K63-polyUb类似于一个“分子桥梁”,促进了MDA5CARDs四聚体的组装,使之形成一个激动状态的构象来招募下游MAVSCARD,以进一步促进MAVSCARD的寡聚和激活(图3)。激活状态下的MDA5可以结合并水解ATP,远程提升CARDs-K63-polyUb10的稳定性以持续激活MAVS。该研究弥补了MDA5通路激活与信号传导研究的空白,进一步揭示了长链,非锚定K63-polyUb在细胞内作为内源性激动剂的免疫学功能,为理解泛素分子多样性在抗RNA病毒天然免疫信号传导与调控中的作用提供了新的线索。
上海药物所博士后宋斌和美国NIH Research Associate陈运为论文第一作者,上海药物所郑杰研究员为论文的通讯作者。该工作得到了新加坡南洋理工大学罗大海教授、吴彬教授,美国Scripps研究所Patrick Griffin教授,上海药物所罗成研究员和张乃霞研究员的大力支持,得到了国家自然科学基金、上海市浦江人才计划等项目的支持。
参考文献:
1.Hu, H. and S.C. Sun, Ubiquitin signaling in immune responses. Cell Res, 2016. 26(4): p. 457-83.
2.Zeng, W., et al., Reconstitution of the RIG-I pathway reveals a signaling role of unanchored polyubiquitin chains in innate immunity. Cell, 2010. 141(2): p. 315-30.
3.Peisley, A., et al., Structural basis for ubiquitin-mediated antiviral signal activation by RIG-I. Nature, 2014. 509(7498): p. 110-4.
4.Jiang, X., et al., Ubiquitin-induced oligomerization of the RNA sensors RIG-I and MDA5 activates antiviral innate immune response. Immunity, 2012. 36(6): p. 959-73.
5.Zheng, J., et al., High-resolution HDX-MS reveals distinct mechanisms of RNA recognition and activation by RIG-I and MDA5. Nucleic Acids Res, 2015. 43(2): p. 1216-30.
6.Zheng, J., et al., HDX-MS reveals dysregulated checkpoints that compromise discrimination against self RNA during RIG-I mediated autoimmunity. Nat Commun, 2018. 9(1): p. 5366.